
The Late Oligocene-Early Pleistocene sporopollen assemblages and paleovegetation succession of core ZkA01 in Hetou town, Leizhou Peninsula of Guangdong Province
-
摘要:
对广东省雷州半岛新生代钻孔ZKA01中的孢粉化石数据进行了系统研究。通过孢粉数据分析,该钻孔自下而上可划分出8个孢粉组合。晚渐新世,该地区植被的垂直分带明显,周围高山有针叶林分布,平原上生长着落叶阔叶与常绿阔叶混交林,气候温凉湿润。早中新世,该地区的植被类型由落叶阔叶与常绿阔叶混交林演变为常绿阔叶林和热带雨林,气候由温暖干旱变为炎热潮湿。晚中新世,该地区的植被类型又演变为山地针叶林和平原上的落叶阔叶与常绿阔叶混交林,气候重新变得温暖湿润。上新世—早更新世,该地区的植被类型为落叶阔叶与常绿阔叶混交林,气温变化波动明显,总体为温凉湿润的气候。孢粉组合所反映的晚渐新世—早更新世气候变化特征与全球气候变化趋势具有较好的可比性。
Abstract:The data of Cenozoic sporopollen fossils obtained from ZKA01 drill hole in Leizhou Peninsula of Guangdong Province were systematically analyzed.Through the analysis of spore-pollen data, 8 sporopollen assemblages can be recognized from this drill hole.In the late Oligocene, the vegetation types in this area reflected the obvious vertical distribution of vegetation; coniferous forests were distributed in the surrounding mountains, and deciduous broad-leaved and evergreen broad-leaved mixed forests grew in the plain.In the early and middle Miocene, the vegetation types in this area evolved from deciduous broad-leaved and evergreen broad-leaved mixed forests into evergreen broad-leaved forests and tropical rainforests.In the late Miocene, the vegetation types in this area evolved into coniferous forests in mountain areas and deciduous broad-leaved and evergreen broad-leaved mixed forests in plain areas.During the Pliocene-Early Pleistocene period, the vegetation types in this area were deciduous broad-leaved and evergreen broad-leaved mixed forests, and the temperature changes fluctuated significantly.The climate change characteristics of Late Oligocene to Early Pleistocene period were reflected by spore-pollen assemblages, which had good comparability with the trend of global climate change.
-
Keywords:
- sporopollen assemblage /
- paleo-vegetation type /
- paleoclimate /
- Cenozoic /
- Leizhou Peninsula
-
致谢: 本文的孢粉分析与研究得到中国地质大学(武汉)喻建新教授的大力协助与指导,在此深表感谢。
-
![]()
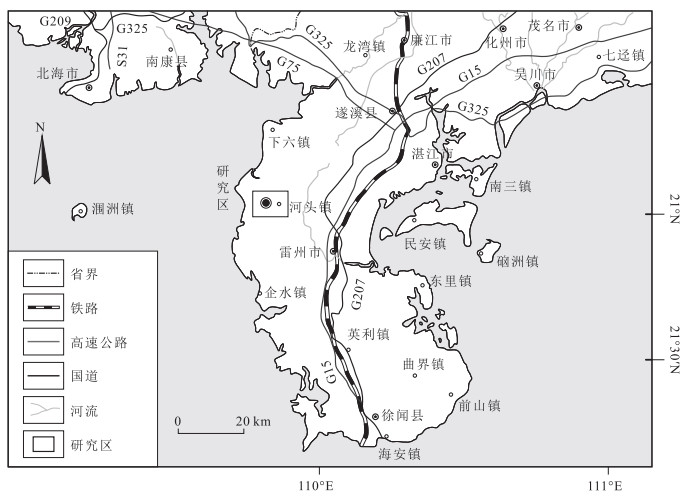
图 1 广东省雷州市河头镇交通位置
Figure 1. Traffic location map of Hetou Town, Leizhou City, Guangdong Province
![]()
图 2 广东省雷州市河头镇地质图
Figure 2. Geological map of Hetou Town, Leizhou City, Guangdong Province
![]()
图 3 广东省雷州半岛ZKA01钻孔渐新世—更新世地层柱状剖面
Figure 3. Columnar section of the Oligocene-Pleistocene strata in ZKA01 drill hole of Leizhou Peninsula, Guangdong Province
![]()
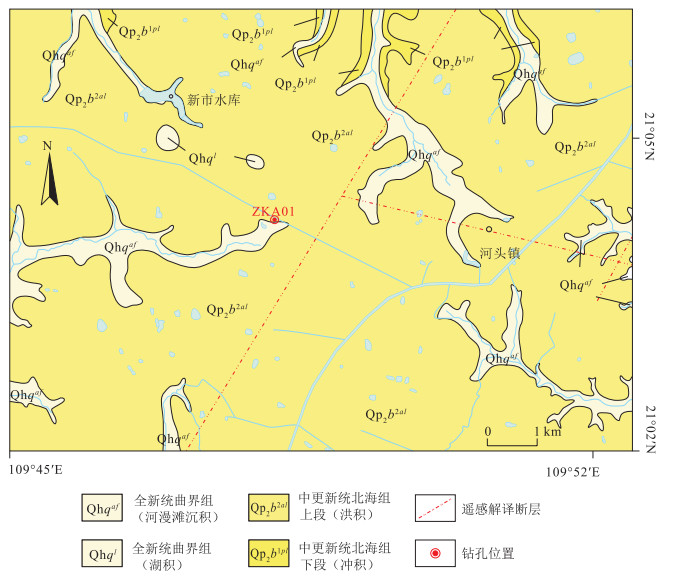
图版Ⅰ
1.Hicriopteris(BF-301,第108层);2、3.Alsophila(BF-236,第87层);4.Osmunda(BF-130,第50层);5~7.Microlepia(BF-60,第27层);8.Lycopodium(BF-479,第168层);9~11.Pteris(BF-126,第49层);12~14.Polypodiodes(BF-130,第50层);15、16.Ceratopteris(BF-727,第264层);17、18.Selaginella(BF-340,第120层);19.Pinus(BF-290,第103层);以上化石照片中的比例尺均为10 μm,化石标本保存于中国地质大学(武汉)地质调查研究院
图版Ⅰ.
![]()
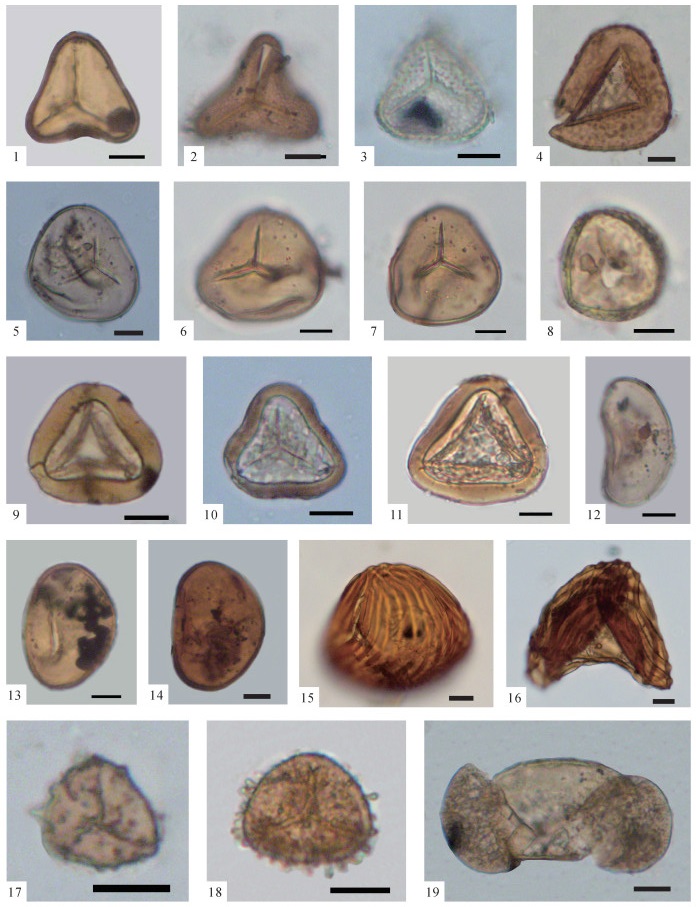
图版Ⅱ
1、2.Pinus(BF-780,第282层);3、4.Tsuga(BF-844,第304层);5、6.Dacrydium(BF-580,第209层);7.Podocarpus(BF-60,第27层);8、9.Ilex(BF-130,第50层);10.Rhus(BF-792,第285层);11、12. Toxicodendron(BF-395,第146层);13.Alnus(BF-780,第282层);14.Betula(BF-344,第122层);15.Corylus(BF-344,第122层);16、17.Chenopodium(BF-276,第95层);18.Aster(BF-205,第84层);以上化石照片中的比例尺均为10 μm,化石标本保存于中国地质大学(武汉)地质调查研究院
图版Ⅱ.
![]()
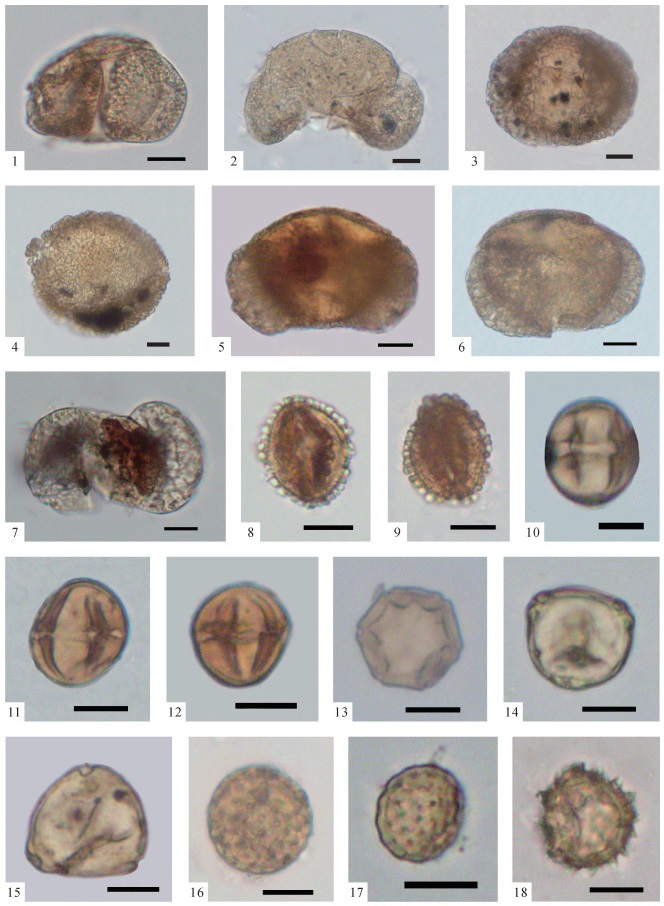
图版Ⅲ
1、2.Euphorbiaceae(BF-826,第293层);3.Rhododendron(BF-236,第87层);4~6.Quercus(BF-130,第50层);7、8.Castanea(BF-271,第95层);9.Lithocarpus(BF-60,第27层);10.Castanopsis(BF-603,第225层);11、12.Cyclobalanopsis(BF-957,第357层);13、14.Gramineae(BF-375,第139层);15~17.Altingia(BF-853,第307层);18.Caesalpinia(BF-313,第113层);19、20.Carya(BF-580,第209层);21、22.Juglans(BF-746,第269层);以上化石照片中的比例尺均为10 μm,化石标本保存于中国地质大学(武汉)地质调查研究院
图版Ⅲ.
![]()
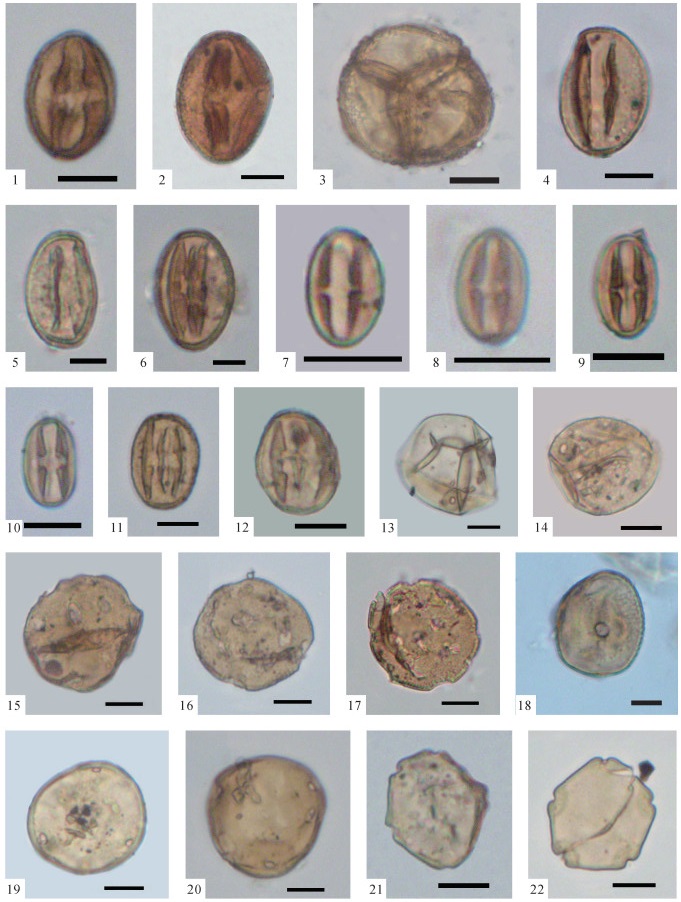
图版Ⅳ
1、2.Platycarya(BF-746,第269层);3.Labiatae(BF-166,第65层);4.Rose(BF-375,第139层);5、6.Myrica(BF-517,第186层);7.Typha(BF-241,第87层);8、9.Polygonum(BF-241,第87层);10、11.Rhodomyrtus(BF-948,第352层);12.Salix(BF-363,第133层);13、14. Calamus(BF-844,第304层)15、16.Potamogeton(BF-727,第264层);17.Symplocos(BF-941,第346层);18.Koelreuteria(BF-236,第87层);19、20.Ulmus(BF-780,第282层);以上化石照片中的比例尺均为10 μm,化石标本保存于中国地质大学(武汉)地质调查研究院
图版Ⅳ.
![]()
图 4 广东省雷州市河头镇ZKA01钻孔渐新世—更新世地层孢粉百分比图谱与孢粉组合划分
Figure 4. Oligocene-Pleistocene spore-pollen percentage diagram and palynological assemblages from ZKA01 drill hole in Hetou Town, Leizhou City, Guangdong Province
表 1 研究区与南海北部大陆边缘盆地新生代地层格架(据参考文献[12]修改)
Table 1 Cenozoic stratigraphic framework of the study area and the northern continental marginal basins of the South China Sea
地质年代 年龄
/Ma珠江口盆地 琼东南盆地 莺歌海盆地 北部湾盆地 雷琼盆地
(本文)纪 世 新近纪 上新世 2.58 琼海组 乐东组 乐东组 湛江组 湛江组 中新世 晚 5.33 万山组 莺歌海组 莺歌海组 望楼港组 望楼港组 中 11.63 粤海组 黄流组 黄流组 灯楼角组 灯楼角组 早 15.97 朝江组 梅山组 梅山组 角尾组 角尾组 古近纪 渐新世 晚 23.03 珠江组 三亚组 三亚组 下洋组 下洋组 28.1 珠海组 陵水组 陵水组 涠洲组 涠洲组  下载: 导出CSV
下载: 导出CSV
-
孙湘君, 孔昭宸, 李明兴.中国南海北部早第三纪花粉新属种[J].植物学报, 1980, 22(2):191-197, 219-220. http://www.cnki.com.cn/Article/CJFD1980-ZWXB198002016.htm 孙湘君, 孔昭宸, 李彭, 等.南海北部早第三纪涠洲组孢粉组合[J].植物分类学报, 1981, 19(2):186-194. 张一勇.雷州半岛第三纪孢粉[J].古生物学报, 1981, 20(5):69-122. http://www.cnki.com.cn/Article/CJFDTotal-GSWX198105007.htm 杨祖年.广东第三系孢粉优势组合带与我国东部各盆地的对比[J].海洋地质与第四纪地质, 1982, 2(4):65-73. http://www.cnki.com.cn/Article/CJFDTotal-HYDZ198204010.htm 童国榜, 龙江平, 李团结, 等.粤西琼东海域表层孢粉沉积的形成环境[J].科学通报, 2014, 59(8):687-700. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=kxtb201408007 张克信, 徐亚东, 骆满生, 等.中国新生代大地构造图[M].北京:地质出版社, 2014. 徐亚东, 梁银平, 江尚松, 等.中国东部新生代沉积盆地演化[J].地球科学——中国地质大学学报, 2014, 39(8):1079-1098. https://www.zhangqiaokeyan.com 张虎男, 赵希涛.雷琼地区新构造运动的特征[J].地质科学, 1984, 19(3):276-287. http://www.cnki.com.cn/Article/CJFDTotal-DZKX198403004.htm 黄玉昆, 邹和平.雷琼新生代断陷盆地构造特征及其演化[J].中山大学学报(自然科学版), 1989, 28(3):1-10. doi: 10.3321/j.issn:0529-6579.1989.03.001 Su M, Zhang C, Xie X N, et al.Controlling factors on the submarine canyon system:A case study of the Central Canyon System in the Qiongdongnan Basin, northern South China Sea[J].Science China:Earth Sciences, 2014, 57(10):2457-2468. doi: 10.1007/s11430-014-4878-4
Su M, Hsiung K H, Zhang C, et al.The linkage between longitudinal sediment routing systems and basin types in the northern South China Sea in perspective of source-to-sink[J].Journal of Asian Earth Sciences, 2015, 111(2015):1-13. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=bb1a267dfe1ec54e5c037542254ecd6b
解习农, 任建业, 王振峰, 等.南海大陆边缘盆地构造演化差异性及其与南海扩张耦合关系[J].地学前缘, 2015, 22(1):77-87. http://d.old.wanfangdata.com.cn/Periodical/dxqy201501007 赵迎冬, 甘华军, 陈善斌, 等, .珠琼运动的厘定——来自北部湾盆地福山凹陷构造沉积特征的启示[J].中国地质, 2015, 42(4):948-959. doi: 10.3969/j.issn.1000-3657.2015.04.012 张克信, 何卫红, 骆满生, 等.中国沉积岩建造与沉积大地构造演化[M].北京:地质出版社, 2017:1-604. 庄文明, 刘建雄, 黄继春, 等.中国区域地质志·广东志[M].北京:地质出版社, 2019. 吴征镒, 孙航, 周浙昆, 等.中国种子植物区系地理[M].北京:科学出版社, 2010:1-485. Zhao Q H, Jian Z M, Wang J L, et al.Neogene oxygen isotopic stratigraphy, ODP Site 1148, northern South China Sea[J].Science in China, 2001, 44(10):934-942. http://d.old.wanfangdata.com.cn/Periodical/zgkx-ed200110010
Wang J, Wang Y J, Liu Z C, et al.Cenozoic environmental evolution of the Qaidam Basin and its implications for the uplift of the Tibetan plateau and the drying of Central Asia[J].Palaeogeogr.Palaeoclimatol.Palaeoecol, 1999, 152:37-47. doi: 10.1016/S0031-0182(99)00038-3
Zachos J, Pagani M, Sloan L, et al.Trends, rhythms, and aberrations in global climate 65 Ma to present[J].Science, 2001, 292:686-693. doi: 10.1126/science.1059412
Holbourn A, Kuhnt W, Kochhann K G D, et al.Global perturbation of the carbon cycle at the onset of the Miocene Climatic Optimum[J].Geology, 2015, 43(2):123-126. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=a1399f7a9453dbf1ffb9961ec1be0c99
Miller K G, Wright J D, Fairbanks R G.Unlocking the Ice House:Oligocene-Miocene oxygen isotopes, eustasy, and margin erosion[J].Journal of Geophysical Research Solid Earth, 1991, 96(B4):6829-6848. doi: 10.1029/90JB02015
Miller K G, Feigenson M D, Kent D V, et al.Upper Eocene to Oligocene isotope(87Sr/86Sr, δ18O, δ13C)standard section, Deep Sea Drilling Project Site 522[J].Paleoceanography, 1988, 3(2):223-233. doi: 10.1029/PA003i002p00223
安芷生, 王苏民, 吴锡浩, 等.中国黄土高原的风积证据:晚新生代北半球大冰期开始及青藏高原的隆升驱动[J].中国科学(D辑), 1998, 28(6):481-490.



计量
- 文章访问数: 2380
- HTML全文浏览量: 368
- PDF下载量: 1757
