
Distribution and influencing factors of soil microbial carbon in plant rhizosphere in a Forest Park, Wuhan
-
摘要:研究目的
土壤微生物生物量碳是土壤有机质中最活跃的组分,研究植物根际土壤微生物量碳对认识土壤碳汇及土壤肥力具有重要意义。
研究方法以武汉市马鞍山森林公园为研究对象,选择4种不同植物类型(乔木、小乔木、灌木、草本)随机设置33个采样点,研究不同植物群落根际土壤微生物量碳分布特征的主要驱动因子。
研究结果①土壤微生物量碳在不同植物群落根际间存在显著差异,根际土壤微生物量碳的波动范围为270.76 ~ 908.44 mg/kg。②土壤微生物量碳与土壤有机碳(r=0.662, p < 0.01)、无机氮(r=0.510, p < 0.01)、碳磷比(r=0.519, p < 0.01)、铵态氮(r=0.355, p < 0.01)和硝态氮(r=0.485, p < 0.01)显著正相关,而与土壤速效磷(r=−0.134,p<0.05)显著负相关。③不同植物群落根际间的土壤微生物生物量熵碳的变化范围为1%~4%,其中黑足鳞毛蕨植物群落的根际最高,桂花植物群落根际最弱。④土壤有机碳、碳磷比和无机碳是影响土壤微生物生物量碳的主要因子,而碳氮比(36.36%,p < 0.01)和有机碳(24.42%,p < 0.05)是决定土壤微生物生物量熵碳含量的关键。
结论土壤碳氮比和有机碳是土壤微生物量熵碳的主要影响因子。不同植物根际土壤中微生物生物量碳间存在显著差异,相比之下黑足鳞毛蕨的根际土壤微生物固碳能力最高。
Abstract:ObjectiveSoil microbial biomass carbon is the most active component of soil organic matter. Studying the microbial biomass carbon in the rhizosphere soil is of great significance.
MethodsThe study took Ma’an Forest Park in Wuhan City as the research object, and four different plant types (trees, small trees, shrubs, and herbs) were selected to randomly set up 33 sampling points to study the main drivers of the distribution characteristics of rhizosphere soil microbial mass carbon in different plant communities.
Results① Soil microbial biomass carbon was significantly different in the plant rhizosphere of different plant communities. The fluctuation range of soil microbial biomass carbon in the plant rhizosphere was 270.76 ~ 908.44 mg/kg. ② Soil microbial biomass carbon was significantly and positively correlated with soil organic carbon (r = 0.662, p < 0.01), inorganic nitrogen (r = 0.510, p < 0.01), carbon−to−phosphorus ratio (r = 0.519, p < 0.01), ammonium nitrogen (r = 0.355, p < 0.01), and nitrate nitrogen (r = 0.485, p < 0.01), while it was significantly and negatively correlated with soil quick−acting phosphorus (r =− 0.134, p < 0.05). ③ The variation of soil microbial biomass entropy carbon in the rhizosphere of different plant communities ranged from 1% to 4%, among which the rhizosphere of the Dryopteris fuscipes C. Chr. was the highest, and that of the osmanthus sp was the lowest. ④ Soil organic carbon, carbon to phosphorus ratio and inorganic carbon were the main factors affecting soil microbial biomass carbon, while carbon to nitrogen ratio (36.36%, p < 0.01) and organic carbon (24.42%, p < 0.05) were the key determinants of entropic carbon content of soil microbial biomass.
ConclusionSoil carbon to nitrogen ratio and organic carbon were the main influencing factors of soil microbial biomass entropy carbon. There were significant differences between the soil microbial biomass carbon in the inter−root soils of different plants, compared to the Dryopteris fuscipes C. Chr, which had the highest inter−root soil microbial carbon sequestration capacity.
-
Keywords:
- soil microbial mass carbon /
- forest soil /
- stoichiometric ratio /
- plant rhizosphere /
- forest park /
- impact factor /
- Wuhan
创新点城市森林有益于缓解城市生态问题,研究环境因子对土壤微生物生物量碳的调控机理有助于土壤碳累积能力,研究结果对城市森林公园碳汇管理和城市可持续发展具有重要的意义。
-
海南岛地热资源非常丰富,按其成因可分为水热型和干热岩型两大类,水热型地热资源又可分为沉积盆地型地热资源和隆起山地型地热资源,隆起山地型地热资源也称构造裂隙型。海南岛隆起山地型地热资源主要分布在中南部丘陵和山地,受导热、导水断裂控制,能量传递方式以热对流为主,热储为呈带状分布的断裂破碎的构造岩。目前,在海南岛中南部已发现隆起山地型地热田16个、温泉30处(陈颖民,2008;高芳蕾等,2009;赵童,2016;张颖,2019),其中,地热田是经过勘查或勘探后证实具有一定规模的地热资源,温泉指出露地表的单个或多个热矿泉,一般勘查程度低,地热资源规模不明确。海南岛隆起山地型地热资源总体开发利用程度较低,仅有10处地热田和温泉得到开发利用,且利用领域单一,主要用于洗浴、旅游酒店,在工业、农业等领域的开发利用基本处于空白。
海南岛隆起山地型地热资源勘查、研究程度差异较大,但总体程度较低。较详细的地热资源调查是1962年由广东省地质局水文工程地质队开展的1∶20万、1∶50万海南岛水文地质概查,对发现的温泉点进行了调查,并提交海南岛热矿泉调查表。1981年由海南地质大队完成的1∶20万海南岛区域水文地质普查,对全岛的地热资源情况进行了初步总结。海南建省后,先后对万宁兴隆、三亚南田、琼海官塘、儋州蓝洋、澄迈西达、保亭七仙岭等9处地热田和13处温泉点进行了勘查评价工作。前人对海南岛隆起山地型地热资源的研究,主要是依据地热田(温泉)勘查评价报告对单个地热田(温泉)的地热地质、水化学特征和地热成因、资源利用和地热勘查方法开展研究(杨兴沐,2004;陈颖民,2008;高芳蕾等,2009;吴小洁,2015;赵童,2016;徐单,2017;杨峰等,2018;张颖,2019)。但对海南岛隆起山地型地热资源形成的地质背景、地热地质特征、资源赋存规律、成因机制等综合研究和评价相对薄弱,在一定程度上制约了海南岛中南部隆起山地型地热资源的潜力认识和开发利用。
本次研究结合区域地质调查、地球物理、水文地质、区域地质调查及地热田(温泉)勘查、调研等资料,开展海南岛隆起山地型地热资源地质综合研究,通过对典型地热田地热地质要素解剖,基本明确了海南岛隆起山地型地热资源地热地质特征和成因地质模型及赋存规律;利用海南岛水文地质勘查钻孔水温资料和地热田、温泉点水温资料,编制了海南岛恒温层水温等值线图,以分析海南岛隆起山地型地热资源分布与恒温层温度的关系;应用前人编制的海南岛居里等温面深度等值线图计算出地温梯度,利用大地热流值定义得到大地热流值,最后用北部福山凹陷油气勘探资料和收集的海南岛46个地热田数据、温泉水温数据、可采储量数据,应用水文地质法估算了地热资源量,评价了海南岛隆起山地型地热资源潜力;依据温度、可采资源规模,分别对海南岛隆起山地型地热田和温泉进行了分类;综合海南岛隆起山地型地热地质特征、赋存规律及资源潜力评价,指出海南岛东南部和西北部是隆起山地型地热资源有利区。
1. 地热地质背景特征
海南岛具有优越的隆起山地型地热资源地质背景,是中国隆起山地型地热资源较丰富的地区。燕山期,海南岛地壳区域性的伸展减薄,地幔物质上涌,深部热载体(岩浆、热液)沿深大断裂带喷溢地表或停留于地壳浅部,从而形成区域性的高热流值(海南省地质调查院,2017)。新生代,海南岛在南北向拉张应力作用下发生持续扩张,形成一系列深大断裂,这些断裂控制了海南岛构造格局、地层、侵入岩、火山岩、地热、地震等分布(海南省地质调查院,2017)。海南岛地热田和温泉均分布于深大断裂附近或断裂交会部位。
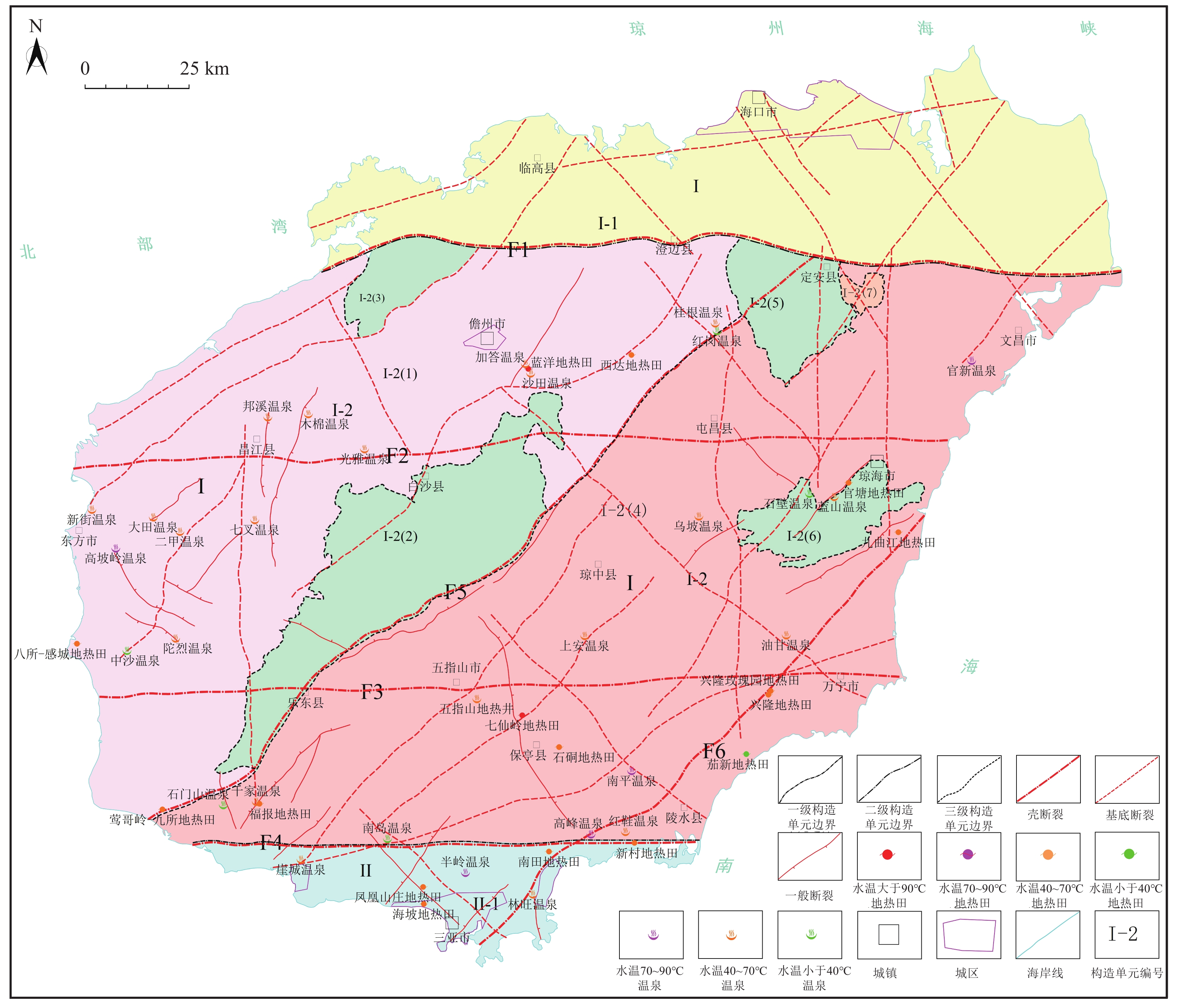
海南岛所处的大地构造位置独特,自20世纪50年代以来,不同学者从不同的大地构造观点、不同的研究目的对海南岛的大地构造单元提出了不同的划分方案(夏邦栋,1991;晁会霞等,2016),尽管认识众说纷纭,但都认为东西向九所-陵水构造带南北两侧古生代在沉积、变质、变形、古生物面貌等方面具有明显的差异性,东西向王五-文教断裂带南北两侧新生代火山作用和沉积作用的差异性,都将这2条东西向断裂带作为一、二级构造单元的边界。参考《中国区域地质表·海南志》(海南省地质调查院,2017)的大地构造划分方案,以东西向九所-陵水断裂带为界,划分出北面的华南褶皱系和南面的南海地台2个一级构造单元,华南褶皱系以王五-文教断裂带为界划分为五指山褶皱带、雷琼断陷2个二级构造单元,五指山褶皱带以北东向的定安-乐东断裂带为界划分为抱板隆起区、五指山隆起区和5个中、新生代断陷成因的凹陷共7个三级构造单元,南海地台只划分为三亚台褶带1个二级构造单元(图1)。海南岛五指山褶皱带的抱板隆起区、五指山隆起区和三亚台缘坳陷3个构造单元,多为中生代侵入岩和古生界变质岩裸露区,岩石具有高的热导率,且深大断裂发育,是大地热流高值异常区。已知的隆起山地型地热田、温泉均分布于这3个构造单元(图1)。
![]() 图 1 海南岛断裂系统、区域构造单元划分与地热田(温泉)分布图(图中断裂分布据海南省地质调查院,2017修改)Ⅰ—华南褶皱系;Ⅱ—南海地台;Ⅰ-1—雷琼断陷;Ⅰ-2—五指山褶皱带;Ⅱ-1 —三亚台缘拗陷带;Ⅰ-2(1)—抱板隆起区;Ⅰ-2(2)—白沙坳陷;Ⅰ-2(3)—王五凹陷;Ⅰ-2(4)—五指山隆起区;Ⅰ-2(5)—雷鸣凹陷;Ⅰ-2(6)—阳江凹陷;Ⅰ-2(7)—长昌坳陷;F1—王五-文教断裂;F2—昌江-琼海断裂;F3—尖峰-吊罗断裂;F4—九所-陵水断裂带;F5—定安-乐东断裂;F6—琼海-三亚断裂Figure 1. Distribution map of fault system, regional tectonic units and geothermal fields (hot springs) in Hainan Island
图 1 海南岛断裂系统、区域构造单元划分与地热田(温泉)分布图(图中断裂分布据海南省地质调查院,2017修改)Ⅰ—华南褶皱系;Ⅱ—南海地台;Ⅰ-1—雷琼断陷;Ⅰ-2—五指山褶皱带;Ⅱ-1 —三亚台缘拗陷带;Ⅰ-2(1)—抱板隆起区;Ⅰ-2(2)—白沙坳陷;Ⅰ-2(3)—王五凹陷;Ⅰ-2(4)—五指山隆起区;Ⅰ-2(5)—雷鸣凹陷;Ⅰ-2(6)—阳江凹陷;Ⅰ-2(7)—长昌坳陷;F1—王五-文教断裂;F2—昌江-琼海断裂;F3—尖峰-吊罗断裂;F4—九所-陵水断裂带;F5—定安-乐东断裂;F6—琼海-三亚断裂Figure 1. Distribution map of fault system, regional tectonic units and geothermal fields (hot springs) in Hainan Island《中国区域地质志·海南志》(海南省地质调查院,2017)利用重、磁异常资料,结合地质调查和火山岩、温泉分布推断的区域断裂有43条,其中东西向5条、南北向4条、北东向18条、北西向16条;按规模和切割深度划分,切割深度达地幔顶部的壳断裂6条、规模较大的切割基底断裂28条、规模较小的一般断裂9条(图1)。海南岛6条壳断裂控制了隆起山地型地热田或温泉的分布,已知的地热田和温泉均分布于东西向王五-文教断裂以南地区,形成3个东西向和2个北东向地热田和温泉带,即沿东西向尖峰-吊罗断裂带、昌江-琼海断裂带、九所-陵水断裂带分布的地热田和温泉带,沿北东向琼海-三亚断裂带和定安-乐东断裂带分布的地热田和温泉带。
海南岛隆起山地型地热田、温泉与中生代花岗岩有密切关系,全岛目前发现的46处隆起山地型地热田、温泉点中有41处直接出露于中生代花岗岩侵入体中,仅二甲温泉出露于中元古代侵入岩中,而新生代火山岩中未发现地热田和温泉出露。其中分布于二叠纪侵入岩中的地热田(温泉)点有11个,分布于三叠纪侵入岩中的地热田(温泉)点有16个,分布于白垩纪侵入岩中的地热田(温泉)点有14个。沉积岩层中温泉和地热田不太发育,仅有九曲江、沙田、蓝洋、桂根4个温泉或地热田分布于石炭系或志留系浅变质砂岩与板岩中。
2. 地热地质特征
本次利用海南岛地球物理、水文地质、区域地质调查、地热田勘查等资料,对海南岛隆起山地型地热田(温泉)地热地质要素进行了系统分析,初步总结了海南岛隆起山地型地热资源的大地热流背景、地温场、储层、导热和导水断层、地热水温、水量、水化学等地热地质特征,并探讨了海南岛隆起山地型地热资源的成因地质模式。
2.1 恒温层温度分布特征
本次收集了海南岛水文地质勘查323口钻孔和已知的46个地热田、温泉点钻孔水温资料(赵童,2016;张颖,2019),编制完成海南岛近地表(相当于恒温层)水温等值线图(图2)。
海南岛东南部和西北部恒温层温度较高,一般大于27℃,中部及北部恒温层温度较低,一般为25~26.5℃。46个地热田、温泉点均位于恒温层温度高异常区。
2.2 居里等温面与大地热流背景
高维等(2020)利用海南岛高精度航磁数据,通过频率域功率谱法反演出居里等温面的深度,海南岛居里等温面的上隆与坳陷大部分呈带状分布,居里等温面存在两隆两坳,即琼中-琼东南隆起区、琼西北隆起区和琼东北坳陷区、乐东-白沙坳陷区。琼中-琼东南隆起区呈北东向分布于琼海、陵水、万宁、保亭、三亚一带,是海南岛居里面埋深最浅的地区,其居里等温面深度为16~24 km,极值点位于陵水北东,深度仅为16 km;琼西北隆起区呈北东向分布于儋州市西、昌江西,至东方市沿海一带,居里等温面深度变化于17~25 km之间;乐东-白沙坳陷区呈北东向分布在海南岛西南部的乐东、中部白沙、儋州市东南、屯昌北一带,居里等温面深度总体变化于26~32 km之间,显示块状沉降区特征,琼东北坳陷区南部边界大致为王五-文教深大断裂带,北至海南岛东北部海岸,对应琼北的新生代断陷区,呈近块状或带状分布,其居里等温面深度为26~35 km,澄迈—临高一带是海南岛居里等温面埋深最大的地区,最大埋深达35 km,在龙州市一带,坳陷带内分布一个北西向次级隆起,居里等温面深度为26 km左右。
大地热流比其他地热单项参数(温度、地温梯度)更能准确地反映一个地区地热场的基本特点,具有十分重要的理论价值和现实作用。本次研究按照大地热流的定义,利用公式(1)计算出海南岛各点的大地热流值。
q=100Kr⋅K (1) 式中:q为大地热流(mW/m2);Kr为岩石导热率(cal/cm·s·℃),其值用全岛分布最广的花岗岩平均热导率3.1 W/(m·oK)(胡圣标等,2001),1 cal/cm·s·℃=418.68 W/m·oK;K为地温梯度(℃/100m)。
地温梯度计算公式如下:
K=(T−t)/H×100 (2) 式中:K为平均地温梯度(℃/100m);T为居里面温度(550℃);t为恒温层温度(℃),由海南岛水文地质勘查323口钻孔水温资料和已知的46个地热田(温泉)点钻孔水温资料编制(图2);H为居里等温面深度(m),按高维等(2020)反演的居里等温面深度取值。
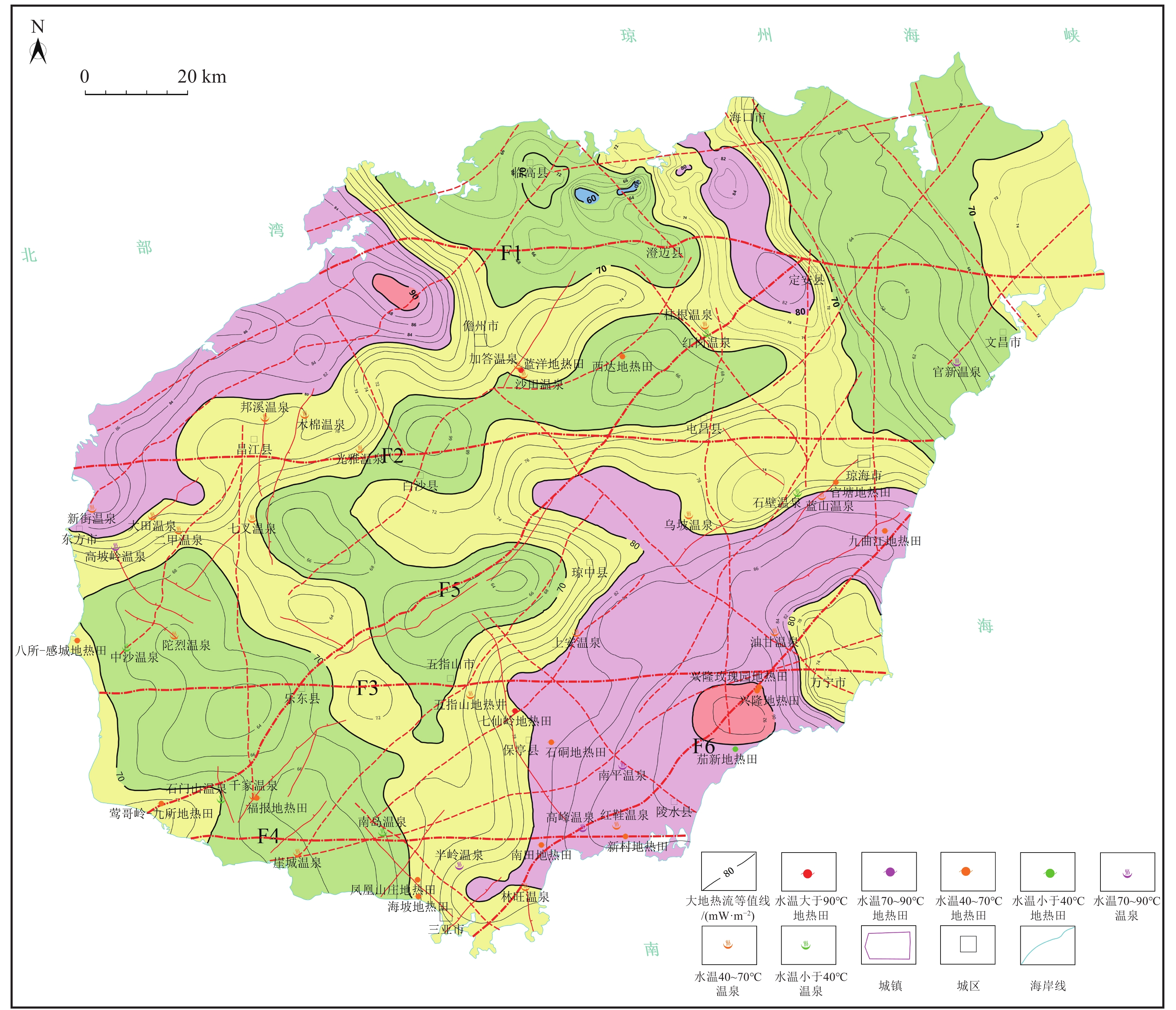
北部福山凹陷的大地热流值根据油气钻井实测6口井的地温梯度与25口井75个岩石样品的实测热导率,计算得到128个热流值点,最后根据前人在海南岛实测的大地热流值(胡圣标等,2001;姜光政等,2016)对计算出的大地热流值进行校正。首次编制完成了海南岛大地热流值等值线图(图3)。
海南岛内大地热流值变化于60~92 mW/m2之间,平均值为76 mW/m2,与全球典型克拉通型大陆区和大洋区的大地热流值相比(高维等,2020;蔺文静等,2016),海南岛总体上具有中等—较高的区域大地热流值。海南岛的大地热流值分布与居里等温面起伏具有较好的相关性,呈北东向或北西向展布(图3)。较高的热流值分布于琼中-琼东南和琼西北2个居里等温面隆起区,琼中-琼东南热流高值区呈北东向展布,位于琼海—万宁—陵水—保亭—三亚一带,大地热流值大于80 mW/m2,最大值在兴隆南,达92 mW/m2,处于东西向尖峰-吊罗断裂与北东向琼海-三亚断裂交会部位;琼西北热流高值区亦呈北东向分布于儋州市西—东方市一带,大地热流值也大于80 mW/m2,最大值在儋州市西,达90 mW/m2,处于东西向王五-文教断裂与北东向博厚-三更断裂交会部位;而居里等温面坳陷区,大地热流值相对较低,如琼北居里等温面坳陷区具有较低的大地热流值,介于60~70 mW/m2之间,最低值区位于福山凹陷中部,与居里等温面埋深等值线一样,在龙州市一带分布1个北西向高值区,大地热流值为80~84 mW/m2,其对应北西向的石山-定安断裂,处于福山凹陷与云龙凸起之间;乐东-白沙居里等温面坳陷区,大地热流值为70~80 mW/m2,变化相对较平缓。
从海南岛大地热流等直线图(图3)可以看出,海南岛已知的46个地热田、温泉点主要分布于东南部和西北部2个大地热流值高值区,说明海南岛隆起山地型地热资源具有高的大地热流背景。
2.3 热储层和热盖层
海南岛隆起山地型地热田、温泉热储层均为断裂破碎的构造岩,控制热储分布的断裂均为张性断层,也是地热田和温泉的导热、导水断层,热储分布于断层破碎带中。热储原岩主要为中生代花岗岩,岩性主要为正长花岗岩、二长花岗岩、花岗闪长岩,只有少数为浅变质沉积岩,岩性为下石炭统南好组石英砂岩、砂岩与板岩和志留系千枚岩、板岩及变质砂岩为原岩的断裂破碎带。
热储层均呈带状构造,为视电阻率低异常带,视电阻率一般小于200 Ω·m,岩石破裂,节理裂隙发育,多具碎裂、碎斑、糜棱、片麻状等结构。热储物性具有强的非均质性,断层上盘物性优于下盘,断层中部优于两侧、浅部优于深部。热储分布范围取决于导热、导水断裂规模,统计官塘、九曲江、蓝洋、七仙岭、南田5个典型地热田热储延伸长度为1.3~3.7 km,宽0.4~1.6 km,面积0.5~2.5 km2,分布于2条断层交会处的热储长宽比较小,单条地层控制的热储长宽比较大,热储埋深不超过500 m。
海南岛隆起山地型地热田热盖层主要为第四系松散沉积层和地表致密花岗岩体。第四系松散沉积层主要为河流冲洪积中粗砂、含砾亚粘土、粘土,其他地区为下伏基岩的坡残积亚粘土或含砾亚粘土,厚0~50 m,除个别砂层外,一般隔水隔热性能较好;地表致密花岗岩体盖层岩性主要为块状花岗岩,岩石裂隙不发育或是闭合性的,或被矿物充填,岩体较完整,具有较好的隔水隔热作用。在热盖层不发育或薄弱处以热水泉排泄,形成温泉。
2.4 导热、导水及控储断层
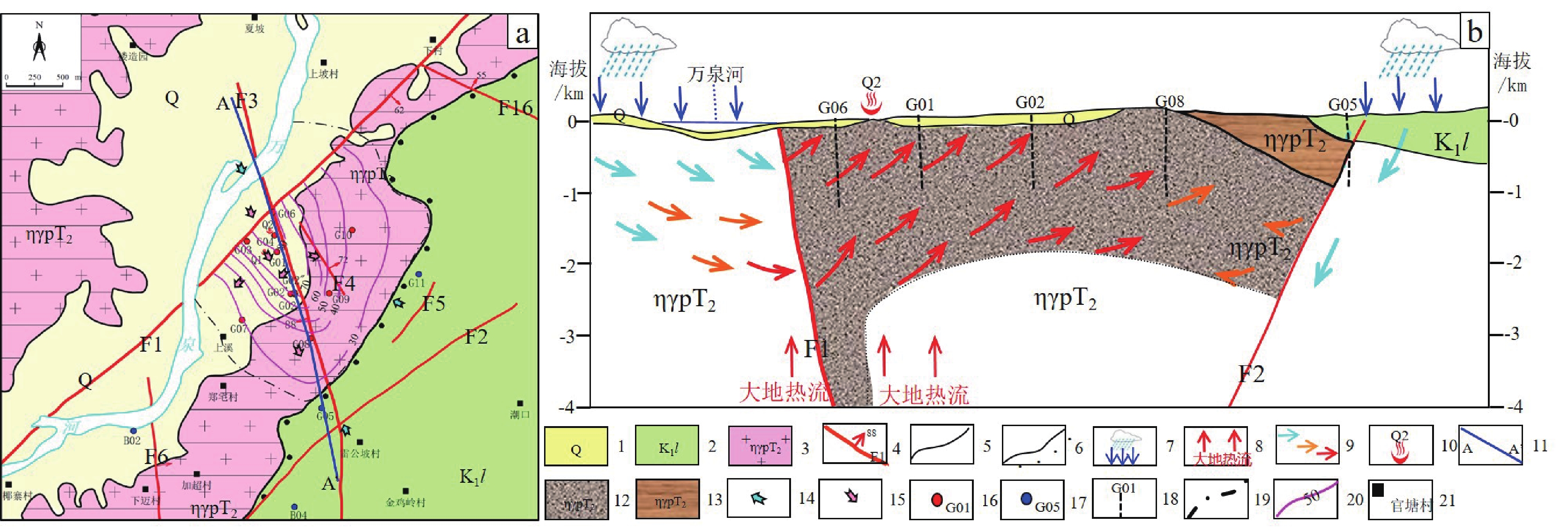
北东向和近东西向基底断裂是形成海南岛隆起山地型地热田和温泉的导热或导水断裂,如官塘地热田的控热断裂为北东向官塘-蓝山断裂,总体走向北东60°,沿万泉河分布,延伸出地热田区外,区内出露长度约为8 km,该断裂是文昌-琼海深大断裂的组成部分,位于其中部;七仙岭地热田导热和导水断层为近东西向,位于东西向尖峰-吊罗断裂南侧,是其旁侧次级断层,出露长度仅2 km左右,地热田温度等值线呈近东西向沿该断层分布,向周边水温降低,说明该断层为七仙岭地热田的导热和导水断层。
海南岛山地隆起型地热资源的导热、导水断裂旁侧的一般断层或更次级断层往往控制地热田(温泉)的热储层分布,从而控制其分布范围和规模,这些断层多为正断层,热储层是受张扭应力作用的断裂破碎带中的构造岩,并且多发育于正断层的上盘。如官塘、蓝洋和南田地热田热储均为北西向张性正断层的上盘破碎带,九曲江地热田热储为北东向张性正断层的上盘破碎带。
2.5 地热水温
海南岛隆起山地型地热田、温泉水温差异较大,地面泉口或井口温度变化范围为35~95℃(赵童,2016;张颖,2019),参考地热资源地质勘查规范(GB/T11615—2010)分类标准,结合海南岛地热资源利用及气候,提出海南岛地热资源温度分类标准(表1)。按照《地热资源地质勘查规范》(GB/T11615—2010)(中华人民共和国国家质量监督检验检疫总局等,2011)分类标准,热水水温为60℃≤t<90℃,考虑海南地区热水水温70℃以上才可用于制冷,将热水下限提高至70℃;海南岛恒温层温度平均在27℃左右,故将水温大于27℃作为地热资源。
表 1 海南岛地热资源按温度分类标准Table 1. Classification standard of geothermal resources by temperature in Hainan Island温度分级 温度(t)区间/℃ 主要用途 GB/T11615-2010 海南岛 高温地热资源 t≥150 t≥150 发电、烘干、采暖 中温地热资源 90≤t<150 90≤t<150 烘干、发电、采暖、制冷 低温地热资源 热水 60≤t<90 70≤t<90 采暖、制冷、理疗、洗浴、温室 温热水 40≤t<60 40≤t<70 理疗、洗浴、采暖、温室、养殖 温水 25≤t<40 27≤t<40 洗浴、温室、养殖、农灌 按表1的地热资源温度分类,海南岛中南部构造裂隙型地热水资源中,中温地热田有七仙岭和蓝洋2个,水温分别为95℃和93℃;低温44个,其中低温热水5个、低温温热水33个、低温温水6个(表2)。
表 2 海南岛地热田、温泉水温统计及分类Table 2. Statistic of water temperature and classification table of geothermal fields and hot springs in Hainan Island序号 地热田、温泉名称 水温/℃ 温度分类 序号 地热田、温泉名称 水温/℃ 温度分类 1 七仙岭地热田 95 中温 24 上安温泉 51 低温温热水 2 蓝洋地热田 93 25 二甲温泉 51 3 高坡岭温泉 78 低温热水 26 兴隆玫瑰园地热田 50 4 南平温泉 77 27 千家温泉 48 5 官新温泉 75 28 木棉温泉 48 6 半岭温泉 77 29 陀烈温泉 47 7 高峰温泉 72 30 七叉温泉 47 8 红鞋温泉 68 低温温热水 31 福报地热田 47 9 官塘地热田 68 32 乌坡温泉 46 10 林旺温泉 67 33 海坡地热田 44 11 兴隆地热田 66 34 新村地热田 42 12 九曲江地热田 64 35 桂根温泉 42 13 南田地热田 60 36 五指山市温泉 40.1 14 崖城温泉 59 37 莺歌岭-九所地热田 40 15 油甘温泉 58 38 光雅温泉 40 16 蓝山温泉 58 39 邦溪温泉 40 17 沙田温泉 57 40 八所-感城地热田 40 18 西达地热田 55 41 中沙温泉 38 低温温水 19 石硐地热田 54 42 红岗温泉 37 20 加答温泉 54 43 茄新地热田 36.1 21 凤凰山庄地热田 53 44 石门山温泉 36 22 大田温泉 52 45 石壁温泉 36 23 新街温泉 51 46 南岛温泉 34.8 海南岛已知的46个地热田、温泉热水水温与控制地热田、温泉的断裂关系密切。一般温度大于70℃的地热田、温泉均分布于海南岛东南部和西北部2个大地热流值高值区;且多分布于6条壳断裂附近。如七仙岭地热田位于东西向尖峰-吊罗断裂带上,官塘、南田等水温较高的地热田位于北东向琼海-三亚断裂上(图1);单个地热田热水温度由地热田中部向边界逐渐降低,导热断层与导水断层交会部位是水温最高的部位,导水断层上盘水温往往高于下盘。如官塘地热田在北东向导热断层与北西向导热断层交会处的泉口水温最高为84℃,向周边水温降低,地热田边界过渡为正常水温,水温大于70℃的地热井均位于北西向导水断层上盘。
2.6 水化学特征及类型
统计海南岛5个典型地热田水质分析资料(赵童,2016;张颖,2019),海南岛隆起山地型地热水化学具有以下特征(表3)。
表 3 海南岛典型地热田地球化学组成Table 3. Summary table of hydrochemical analysis of typical geothermal fields in Hainan Island地热田
名称阳离子/(mg·L−1) 阴离子/(mg·L−1) pH值 矿化度/(g·L−1) 水化学类型 K+ Na+ Ca2+ Mg2+ Fe3+ Cl− SO42− HCO3− SiO42− NO3− F− 七仙岭 2.8 67.2 2.8 0.07 <0.02 12.0 25.9 58.6 128 0.94 12.0 8.98 0.299 HCO3−Na 蓝洋 5.7 84.4 19.2 1.8 <0.02 23.6 66.1 134 151 0.29 14.0 8.32 0.467 HCO3•SO4−Na 官塘 7.9 185 8.8 0.34 <0.02 115 73.8 164 113 0.50 24.0 8.39 0.678 HCO3•Cl−Na 九曲江 58.7 1590 664 4.5 1.0 3410 292 73.2 126 17.3 2.4 7.47 6.21 Cl−Na•Ca 南田 16.0 393 190 2.6 <0.02 741 255 48.8 100 1.7 4.8 7.89 1.73 Cl−Na•Ca (1)阳离子以Na+、Ca2+、K+为主,Na+含量25.1~1590 mg/L,Ca2+含量2.4~664 mg/L,K+含量2.8~58.7 mg/L。九曲江和南田地热田Na+、Ca2+含量高,是其他3个地热田的几十倍。
(2)阴离子以Cl−、SO42−、HCO3−为主,Cl−含量12~3410 mg/L,SO42−含量25.9~292 mg/L,HCO3−含量48.8~164 mg/L。九曲江和南田地热田Cl−含量高出其他3个地热田几十倍。
(3)水化学类型较复杂,变化较大,主要有HCO3−Na、HCO3•SO4−Na、HCO3•Cl−Na、Cl−Na•Ca、HCO3−Na•Ca等类型,靠近海边多为Cl−Na•Ca型,七仙岭地热田属HCO3−Na型,蓝洋地热田属HCO3•SO4−Na型,官塘地热田属HCO3•Cl−Na型,九曲江和南田地热田属Cl−Na•Ca型(图4;表3)。
![]() 图 4 海南岛典型地热田水化学类型Piper图Figure 4. Piper diagram of hydrochemical types in typical geothermal fields in Hainan Island
图 4 海南岛典型地热田水化学类型Piper图Figure 4. Piper diagram of hydrochemical types in typical geothermal fields in Hainan Island(4)热水中F−含量较高,含量为2.4~24 mg/L,达到氟水水质标准,具有医疗价值。
(5)矿化度除九曲江和南田地热田较高外,其他均较低。九曲江地热田矿化度达6.2 g/L,南田地热田为1.73 g/L,是由于热矿水在径流过程中混入海水的原因。
(6)pH值介于7~9之间,为碱性热矿水。
(7)九曲江和南田地热田Na+、Ca2+、Cl−含量高异常及高矿化度的原因是其位于海边,地热水补给除大气降水外,还有海水补给,或热矿水在径流过程中混入海水。
(8)七仙岭、蓝洋、官塘3个远离海边的地热田的Na+、Cl−比值为1.61~5.60,大于海水平均值(0.85),说明地热水为大气成因的渗透水。而九曲江、南田地热田的Na+、Cl−比值分别为0.45和0.53,小于海水平均值,也说明地热水有海水的补给(张玉良等,2019)。
2.7 地热成因模式
高芳蕾等(2009)、赵童(2016)、张颖(2019)利用海南岛地热田和温泉热水同位素δD和δ18O数据进行分析,δD和δ18O均为负值,数据点落在大气降水线上,结果表明,地热田和温泉热水的补给主要为大气降水。热水地球化学分析和水化学类型结果表明,临海的地热田和温泉有海水的补给(表3);根据海南岛典型地热田放射性同位素分析,热水放射性元素含量低(表4),由其所致的热流增量小,因此区内地热田和温泉热源主要来源于地壳深部或地幔的热能;根据海南岛典型地热田热水地球化学分析资料,应用钾镁温标估算的热储温度在74~118℃之间,钾钠温标估算的热储温度在171~202℃之间(表5)。一般认为,钾镁温标公式代表不太深处热储层中的热动力平衡条件,估算的温度为浅层热储温度,而钾钠温标公式是根据水岩平衡和热动力方程推导的计算公式,适用于估算深部热储温度,在略高于海南岛平均地温梯度(3.5℃/100m)的大地热流背景下、恒温层温度取27℃,得到海南岛典型地热田和温泉热储埋深深度为4~5 km(表4)。
表 4 海南岛典型地热田放射性同位素分析及热水循环深度估算Table 4. Radioisotope analysis and the hot water circulation depth estimation of typical geothermal fields in Hainan Island地热田名称 放射性同位素/(Bq·L−1) 阳离子/(mg·L−1) 热储温度/℃ 热水循环深度估算/km 226Ra/10−3 220Ra 总α 总β K+ Na+ Mg2+ K−Mg温标 K−Na温标 七仙岭 2.8 67.2 0.07 99 171 4.11 蓝洋 14.2 17.78 0.512 0.334 5.7 84.4 1.8 76 203 5.02 官塘 4.9 4.65 0.088 0.414 7.9 185 0.34 106 172 4.16 九曲江 23.42 39.91 0.059 0.073 58.7 1590 4.5 127 164 3.90 南田 78.2 50.57 0.58 0.615 16.0 393 2.6 97 169 4.07 表 5 海南岛地热田资源量估算及规模分类Table 5. Resources estimation and classification of geothermal fields in Hainan Island序号 地热田名称 水温/℃ 可采热水储量/(m3·d−1) 可采资源量/(108kcal·d−1) 可采资源量折合标煤/t 热能/MW 规模分类 1 蓝洋地热田 93 7000 4.4740 63.91 21.680 中型 2 西达地热田(含九乐宫温泉) 55 1200 0.3312 4.73 1.605 小型 3 官塘地热田 68 6700 2.6849 38.36 13.011 中型 4 九曲江地热田 64 7800 2.8297 40.42 13.712 中型 5 兴隆地热田 66 7708 2.9475 42.11 14.283 中型 6 兴隆玫瑰园地热田 50 2367.36 0.5380 7.69 2.607 小型 7 茄新地热田 36.1 5774.98 0.5224 7.46 2.532 小型 8 新村地热田 42 2265.28 0.3371 4.82 1.634 小型 9 七仙岭地热田 95 4040 2.6420 37.74 12.803 中型 10 石硐地热田 54 1032 0.2747 3.92 1.331 小型 11 南田地热田 60 8030 2.6051 37.22 12.624 中型 12 凤凰山庄地热田 53 4640 1.1891 16.99 5.762 小型 13 海坡地热田 44 2972.5 0.5004 7.15 2.425 小型 14 福报地热田 47 1385.47 0.2740 3.91 1.328 小型 15 莺歌岭-九所地热田 40 3934 0.5074 7.25 2.459 小型 16 八所-感城地热田 40 2887.48 0.3724 5.32 1.805 小型 合计 6.97×104 23.03 329 111.6 综合海南岛已发现的地热田和温泉地热地质特征,可以将区内地热资源的成因模式归纳为:热源主要来源于地壳深部或地幔的热能,大气降雨通过断裂在地壳深部形成热对流循环,水体得到能量,水温升高,水压增大,使热流体通过裂隙上升,于构造岩体破碎段富集为热矿水体。热矿水一部分沿断裂垂直向上流动,在盖层薄弱处以热水泉排泄,形成温泉;另一部分在热矿泉高水位区向四周作水平流动,到热储边界与地表常水混合,水温降低。热储主要为以花岗岩或变质岩为原岩的断裂破碎带,盖层为第四系松散层或较完整的岩体(图5)。而邻海的地热田或温泉由于海水的侵入,与热水混合,则形成高矿化度、高盐度的咸热水,如九曲江地热田和南田地热田。
![]() 图 5 官塘地热田地热地质图(a)和成因模式图(b)1—第四系;2—白垩系下统鹿母湾组;3—三叠纪中期花岗岩;4—断层;5—地质界线;6—不整合界线;7—地下水补给;8—大地热对流;9—地下水流向;10—地面温泉及编号;11—模式图剖面位置;12—三叠纪中期花岗岩断裂破碎带热储;13—三叠纪中期花岗岩基岩盖层;14—地下水补给方向;15—地热水排泄方向;16—热水钻孔及编号;17—常温钻孔及编号;18—地热钻孔及编号;19—地热田边界;20—水温等值线;21—村镇Figure 5. Geothermal geological map (a) and genetic model ideograph of Guantang geothermal field (b)
图 5 官塘地热田地热地质图(a)和成因模式图(b)1—第四系;2—白垩系下统鹿母湾组;3—三叠纪中期花岗岩;4—断层;5—地质界线;6—不整合界线;7—地下水补给;8—大地热对流;9—地下水流向;10—地面温泉及编号;11—模式图剖面位置;12—三叠纪中期花岗岩断裂破碎带热储;13—三叠纪中期花岗岩基岩盖层;14—地下水补给方向;15—地热水排泄方向;16—热水钻孔及编号;17—常温钻孔及编号;18—地热钻孔及编号;19—地热田边界;20—水温等值线;21—村镇Figure 5. Geothermal geological map (a) and genetic model ideograph of Guantang geothermal field (b)3. 地热资源潜力评价及赋存规律
3.1 地热资源潜力评价
将海南岛已发现或探明的46个隆起山地型地热田和温泉依据水温、可采储量(赵童,2016),利用水文地质法估算可采资源量,计算公式为:
QR1=ρwCwQ(tr−tj) (3) 式中:QR1=QR/t,为单位时间的地热资源量(kcal/d);Q=Adn/t,为热矿水可采资源量(L/d);t为时间(d);ρw为热水密度(kg/m3),查表得到;Cw为水的比热容(1 kcal/kg•℃);tr为热矿水水温(℃);tj为年平均气温,海南岛取27℃。
由于地热田和温泉在勘探程度、可采水量存在巨大差异,一般同一处温泉经勘探后,用地热井开采的可采水量是温泉的10倍左右,故将地热田与温泉分开估算。
将热能单位1 kcal/d换算成MW的换算关系是:
1 W=3.6 kj/h=3.6/4.1868 kcal/h=0.8598×24 kcal/d=20.64 kcal/d。所以,1 MW=20.64×106 kcal/d。
通过估算,海南岛已发现或探明的46个隆起山地型地热田和温泉的总可采热水量约为8.1×104 m3/d,可采资源量为26.17×108 kcal/d,折合126.81 MW,相当于标煤373.83 t/d(表5、表6)。其中,16个地热田可采热水量约为6.97×104m3/d,可采资源量为23.03×108 kcal/d,折合111.6 MW,相当于标煤329 t/d(表5);30个温泉可采热水量约为1.15×104 m3/d,可采资源量为3.14×108 kcal/d,折合15.21 MW,相当于标煤44.84 t/d(表6)。
表 6 海南岛温泉资源量估算及规模分类Table 6. Resources estimation and classification of hot springs in Hainan Island序号 地热田名称 水温/℃ 可采热水储量/(m3·d−1) 可采资源量/(108kcal·d−1) 可采资源量折合标煤/t 热能/MW 规模分类 1 沙田温泉 57 541.44 0.1598 2.28 0.775 小型 2 加答温泉 54 70.85 0.0189 0.27 0.091 小型 3 桂根温泉 42 216 0.0321 0.46 0.156 小型 4 红岗温泉 37 603.36 0.0600 0.86 0.291 小型 5 官新温泉 75 296 0.1385 1.98 0.671 小型 6 蓝山温泉 58 25.92 0.0079 0.11 0.038 小型 7 石壁温泉 36 61.92 0.0055 0.08 0.027 小型 8 乌坡温泉 46 494.4 0.0930 1.33 0.451 小型 9 上安温泉 51 463.68 0.1099 1.57 0.533 小型 10 油甘温泉 58 26 0.0079 0.11 0.038 小型 11 南平温泉 77 1228.8 0.5976 8.54 2.896 中型 12 红鞋温泉 68 174.53 0.0700 1.00 0.339 小型 13 高峰温泉 72 401.76 0.1768 2.53 0.857 小型 14 林旺温泉 67 1054 0.4134 5.91 2.003 中型 15 五指山市地热点 40.1 384 0.0499 0.71 0.242 小型 16 半岭温泉 77 563.24 0.2742 3.92 1.329 中型 17 南岛温泉 34.8 850.18 0.0659 0.94 0.319 小型 18 崖城温泉 59 157.92 0.0497 0.71 0.241 小型 19 千家温泉 48 426 0.0885 1.26 0.429 小型 20 石门山温泉 36 1382 0.1236 1.77 0.599 小型 21 中沙温泉 38 25.92 0.0028 0.04 0.014 小型 22 陀烈温泉 47 75.12 0.0149 0.21 0.072 小型 23 高坡岭温泉 78 667.9 0.3318 4.74 1.608 中型 24 二甲温泉 51 129.6 0.0307 0.44 0.149 小型 25 大田温泉 52 254.02 0.0627 0.90 0.304 小型 26 新街温泉 51 174.53 0.0414 0.59 0.201 小型 27 七叉温泉 47 157.25 0.0311 0.44 0.151 小型 28 邦溪温泉 40 256.56 0.0331 0.47 0.160 小型 29 木棉温泉 48 38.88 0.0081 0.12 0.039 小型 30 光雅温泉 40 298.94 0.0386 0.55 0.187 小型 合计 1.15×104 3.14 44.84 15.21 根据《地热资源地质勘查规范》(GB/T11615—2010)(中华人民共和国国家质量监督检验检疫总局等,2011)分类标准(表7),按同级地热田是温泉热能的10倍,将海南岛中南部构造裂隙型地热水资源划分为中型、小型2类地热资源。其中,中型地热田6个(可采能量大于10 MW),分别为蓝洋、官塘、九曲江、兴隆、七仙岭、南田地热田,小型地热田10个(可采能量小于10 MW);中型温泉4个(可采能量大于1 MW),分别为南平、林旺、半坡、高坡岭温泉,小型温泉26个(可采能量小于1 MW)。
表 7 水热型中低温地热田和温泉规模分类标准Table 7. Classification standards of low temperature geothermal fields and hot springs地热资源
规模地热田分类 温泉分类 热能/MW 保证开采
年限/年热能/MW 保证开采
年限/年大型 > 50 100 >5 100 中型 10∼50 100 1∼5 100 小型 < 10 100 < 1 100 注:地热田规模分类标准按《地热资源地质勘查规范》(GB/T11615—2010);温泉规模分类标准按同级地热田的1/10 3.2 地热资源赋存规律及勘探开发有利目标区
通过对海南岛已知的46个地热田和温泉地热地质综合研究,尤其是对其中5个典型地热田的地质要素进行系统分析,基本明确了海南岛隆起山地型地热资源的赋存规律。
(1)海南岛隆起山地型地热资源均分布于中南部五指山褶皱带的抱板隆起区、五指山隆起区和三亚台缘坳陷3个构造单元。
(2)断裂是控制区内地热田和温泉最主要的地质要素,东西向、北东向深大断裂是主要导热、导水断裂,旁侧的次级断层往往控制地热田(温泉)的热储层分布,热储均为断层破碎带构造岩。
(3)海南岛地热田、温泉多分布于中生代花岗岩区,受断裂破碎的花岗岩是最主要的热储。
(4)海南岛隆起山地型水温高、规模较大的地热田、温泉主要分布于东南部和西北部2个大地热流值高值区。
(5)海南岛隆起山地型地热田、温泉主要分布在恒温层水温高值区。
因此,海南岛东南部和西北部深大断裂附近及其交会部位、大地热流高值区、中生代花岗岩侵入体出露(或隐伏)区、具有高的地面水温的地区是隆起山地型地热资源勘探开发的有利目标区,可能也是隆起山地型干热岩地热资源的有利区。已知地热田及温泉周边是地热资源勘探开发的有利目标区,如官塘地热田F6、F16两条北西向断裂与F1断裂交会部位具有与官塘地热田相似的地热地质条件,是官塘地热田周边寻找隆起山地型地热田的有利目标区(图5−a);七仙岭地热田水温高,地热田深部热储温度可达170℃以上,具备深层水热型和干热岩地热资源勘探开发潜力。
4. 结 论
(1)海南岛隆起山地型地热资源的主要控制因素是东西向和北东向深大断裂,深大断裂是导热、导水断裂,旁侧的次级断层控制热储分布,受断裂破碎的花岗岩是最主要的热储。
(2)海南岛6条壳断裂控制了隆起山地型地热田或温泉的分布,已知的地热田和温泉均分布于东西向王五-文教断裂以南地区,形成沿尖峰-吊罗、昌江-琼海、九所-陵水断裂带分布的3个东西向地热田(温泉)带和沿琼海-三亚、定安-乐东断裂带分布的2个北东向地热田(温泉)带。
(3)海南岛隆起山地型地热资源的热源主要来源于地壳深部或地幔的热能,热水补给为大气降水,通过断裂在地壳深部形成热对流循环,于构造岩体破碎段富集为热矿水体,在盖层薄弱处以热水泉排泄,形成温泉。邻海的地热田或温泉由于海水的侵入,与热水混合,形成高矿化度、高盐度的咸热水。
(4)海南岛隆起山地型地热资源丰富,已发现和探明的46个地热田和温泉可采热水量约8.1×104 m3/d,可采资源量26.17×108 kcal/d,折合126.81 MW,相当于标煤373.83 t/d,按地热田和温泉水温、规模分类属于中、低温和中、小型地热田或温泉。海南岛东南部和西北部深大断裂附近及交会部位、大地热流高值区、中生代花岗岩侵入体出露(或隐伏)区、具有高的恒温层水温的地区是隆起山地型地热资源勘探开发有利区。
蓝洋和七仙岭地热田水温高,目前利用程度较低且单一,热能盈余多,建议加强地热资源用于制冷的可行性研究;海南岛东南部具有高的大地热流背景,主要为中生代花岗岩出露区,分布有七仙岭地热田和南平、高峰等温度较高的温泉,具有干热岩地热资源潜力,建议加大干热岩地热资源研究评价和勘探工作。
-
![]()
图 2 马鞍山森林不同植物群落的土壤微生物生物量碳含量(不同小写字母代表差异显著(p < 0.05)。黑鳞蕨: 黑足鳞毛蕨)
Figure 2. Soil microbial biomass carbon content in different species communities in Ma'an forest
![]()
图 3 马鞍山森林公园土壤微生物生物量熵碳
注:不同小写字母代表差异显著(p < 0.05)。黑鳞蕨: 黑足鳞毛蕨
Figure 3. Soil Microbial Biomass Entropy Carbon in Ma'an Forest Park
![]()
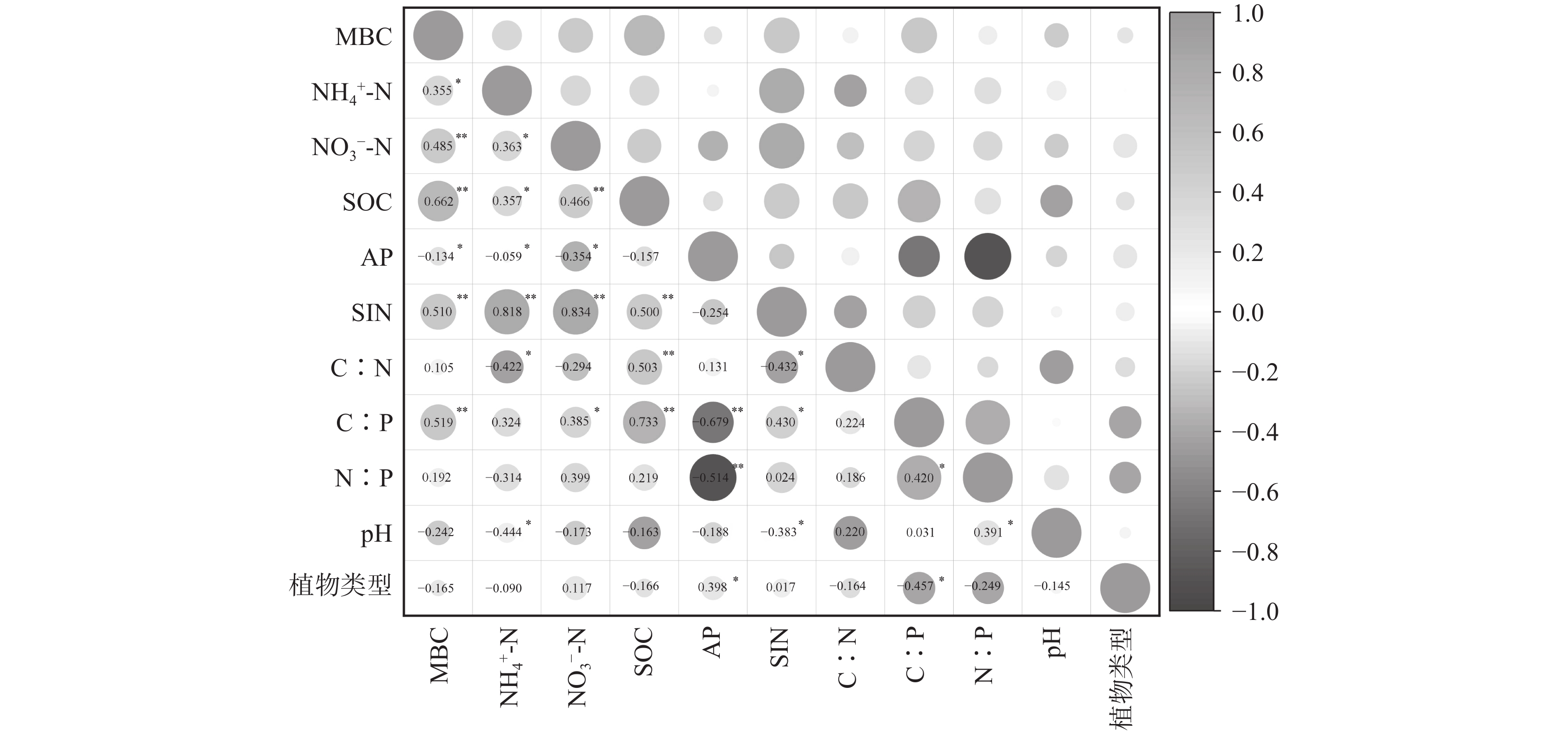
图 4 马鞍山森林公园土壤因子的相关性热图(*表示在0.05级别(双尾)相关性显著, **表示在0.01级别(双尾)相关性显著)
Figure 4. Correlation heat map of Soil Factors in the Ma'an Forest Park
表 1 马鞍山森林土壤理化指标
Table 1 Soil physicochemical indexes of Ma'an Forest Park
物种 有机碳/( g·kg−1) 速效磷/( mg·kg−1) 铵态氮/( mg·kg−1) 硝态氮/( mg·kg−1) 无机氮/( mg·kg−1) pH值 MBC/( mg·kg−1) 乔木 香樟 46.02±4.67a 35.34±1.54c 15.33±1.77a 9.00±1.36a 24.33±1.70a 4.49±0.11c 908.44±163.16a 马尾松 24.21±3.23bc 49.38±3.17b 7.20±0.41c 5.83±0.62bc 13.03±0.36b 4.56±0.08bc 601.69±93.09ab 柏木 23.26±4.10bc 31.82±3.87c 12.53±1.61ab 6.4±1.56ab 18.93±3.07ab 4.78±0.03ab 403.02±112.73b 小乔木 杉松 23.14±1.47bc 50.72±1.88b 13.23±1.36ab 7.87±1.32ab 21.1±2.26ab 4.83±0.13ab 565.60±54.31ab 桂花 20.23±2.69bc 96.90±4.24a 9.7±0.63bc 1.07±0.39bc 10.77±1.03bc 4.56±0.12bc 270.76±77.03b 广玉兰 12.37±1.37c 103.16±2.61a 10.77±1.14bc 0.67±0.11c 11.43±1.03bc 4.91±0.06a 392.62±86.02b 灌木 野鸦椿 31.16±0.76ab 91.66±5.73a 8.70±1.09bc 6.93±1.32a 15.63±0.33b 4.49±0.04bc 676.44±48.12ab 荚蒾 35.51±3.20a 99.21±3.19a 17.90±2.98a 7.20±2.15a 25.10±5.08a 4.38±0.09c 625.69±31.55ab 海桐 30.90±3.70ab 84.25±4.17a 8.33±1.08bc 9.07±1.28a 17.90±1.43ab 4.38±0.11c 681.51±29.41ab 草本 野青茅 23.54±2.95bc 40.55±1.14bc 10.6±0.71bc 9.17±3.31a 19.77±4.02ab 4.79±0.08ab 412.18±98.78b 黑足鳞毛蕨 22.07±3.65bc 51.45±7.93b 12.17±2.21ab 8.63±2.84a 20.80±4.93ab 4.66±0.03ab 514.49±90.73ab 注:不同小写字母代表差异显著(p < 0.05)  下载: 导出CSV
下载: 导出CSV
表 2 马鞍山森林土壤养分化学计量比
Table 2 Stoichiometric ratio of soil nutrient content in Ma'an forest
物种 C∶P C∶N N∶P 乔木 香樟 1311.10±183.87a 1879.87±60.54a 0.75±0.04a 马尾松 522.72±79.36bc 1841.26±204.50a 0.45±0.04bc 柏木 791.97±99.28b 1271.08±177.38ab 0.74±0.07a 小乔木 杉松 453.25±42.37cd 1112.92±50.02b 0.49±0.04bc 桂花 214.75±25.18e 1883.67±222.85a 0.24±0.01d 广玉兰 121.23±11.92e 1114.99±183.82b 0.20±0.004d 灌木 野鸦椿 343.53±21.11de 1998.98±88.47a 0.17±0.01d 荚蒾 376.94±64.68de 1675.95±44.77a 0.26±0.06d 海桐 375.57±61.26de 1718.27±12.12a 0.22±0.03d 草本 野青茅 569.11±73.79bc 1283.25±209.05ab 0.57±0.03b 黑足鳞毛蕨 358.95±48.25de 1118.38±304.52b 0.39±0.06c 注:不同小写字母代表差异显著(p < 0.05)
下载: 导出CSV
-
Bhople P, Djukic I, Keiblinger K, et al. 2019. Variations in soil and microbial biomass C, N and fungal biomass ergosterol along elevation and depth gradients in Alpine ecosystems[J]. Geoderma, 345: 93−103. doi: 10.1016/j.geoderma.2019.03.022
Brookes P C, Landman A, Pruden G, et al. 1985. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil[J]. Soil Biology and Biochemistry, 17(6): 837−842. doi: 10.1016/0038-0717(85)90144-0
Brookes P C, Powlson D S, Jenkinson D S. 1982. Measurement of microbial biomass phosphorus in soil[J]. Soil Biology and Biochemistry, 14(4): 319−329. doi: 10.1016/0038-0717(82)90001-3
Chen M K, Wang S J, Chen W Q, et al. 2019. Effects of ant nesting on soil microbial biomass carbon and quotient in tropical forest of Xishuangbanna[J]. Chinese Journal of Applied Ecology, 30: 2973−2982 (in Chinese with English abstract).
Chen X H, Chen Z Z, Lei J R, et al. 2021. Soil microbial biomass carbon, nitrogen and nutrient characteristics of different plant communities in Dongzhai Port[J]. Forest Resources Management, (6): 97−104 (in Chinese with English abstract).
Cheng Y, Zhang J B, Cai Z C. 2012. A research progress on biotic and abiotic inorganic N immobilization in soils[J]. Acta Pedologica Sinica, 49(5): 1030−1036 (in Chinese with English abstract).
Ding L Z, Man X L, Xiao R H, et al. 2019. Dynamics of soil microbial biomass carbon and nitrogen in the soil of rhizosphere during growing season in the cold temperate forests[J]. Scientia Silvae Sinicae, 55(7): 178−186 (in Chinese with English abstract).
Gao D, Bai E, Wang S, et al. 2022. Three−dimensional mapping of carbon, nitrogen, and phosphorus in soil microbial biomass and their stoichiometry at the global scale[J]. Global Change Biology, 28(22): 1−13.
Gao S, Yin H, Fu M J, et al. 2018. Effects of freeze−thaw cycles on soil microbial biomass carbon, nitrogen, and nitrogen mineralization in three types of forest in the temperate zone[J]. Acta Ecologica Sinica, 38(21): 7859−7869 (in Chinese with English abstract).
Guan H Y, Wang Q, Zhao X, et al. 2015. Seasonal patterns of soil microbial biomass C and impacting factors in two typical arid desert vegetation regions[J]. Arid Land Geography, 38(1): 67−74 (in Chinese with English abstract).
Hu Z D, Liu S R, Shi Z M, et al. 2012. Variations of soil nitrogen and microbial biomass carbon and Nitrogen of Quercus aquifolioides forest at different attitudes in Balangshan, Sichuan[J]. Forest Research, 25(3): 261−268 (in Chinese with English abstract).
Lai J, Zou Y, Zhang J, et al. 2022. Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca. hp R package[J]. Methods in Ecology and Evolution, 13: 782−788. doi: 10.1111/2041-210X.13800
Li P, Muledeer T, Tian D, et al. 2019. Seasonal dynamics of soil microbial biomass carbon, nitrogen and phosphorus stoichiometry across global forest ecosystems[J]. Chinese Journal of Plant Ecology, 43(6): 532−542. doi: 10.17521/cjpe.2019.0075
Li R B. 2008. Review of research advances of soil microbial biomass carbon[J]. Forestry and Environmental Science, 24: 65−68 (in Chinese with English abstract).
Li Y, Lu J. 2023. Factors influencing soil carbon and the impact of litter on soil carbon storage[J]. Agriculture and technology, 43(4): 52−55 (in Chinese with English abstract).
Liu C, Tian J, Cheng K, et al. 2023. Topsoil microbial biomass carbon pool and the microbial quotient under distinct land−use types across China: A data synthesis[J]. Soil Science and Environment, 2(5): 1−10.
Liu S. 2010. Temporal and spatial variations of microbial biomass carbon and nitrogen in five temperate forest soils[D]. Master Thesis of Northeast Forestry University (in Chinese with English abstract).
Ma C X, Li H J, Zhen H F, et al. 2019. Responses of soil active carbon and nitrogen to short−term warming in alpine treeline of west Sichuan, China[J]. Chinese Journal of Applied Ecology, 30(3): 718−726 (in Chinese with English abstract).
Monika R, Kusum A, Ayyanadar A. 2021. Seasonal dynamics in soil microbial biomass C, N and P in a temperate forest ecosystem of Uttarakhand, India[J]. Tropical Ecology, 62(3): 377−385.
Office of the National Soil Census. 1979. Interim technical regulations for the Second National Soil Survey[M]. China Agriculture Press: 1−79 (in Chinese).
Qing Y, Sun F D, Li Y, et al. 2015. Analysis of soil carbon, nitrogen and phosphorus in degraded alpine wetland, Zoige, southwest China[J]. Acta Prataculturae Sinica, 24(3): 38−47 (in Chinese with English abstract).
Shi F J, Huang Z Y, Li T, et al. 2018. Vertical changes of the soil microbial biomass and the correlation analysis in Parashorea chinensis natural forest[J]. Forestry and Environmental Science, 34: 72−76 (in Chinese with English abstract).
Su F, Xu S, Sayer E J, et al. 2021. Distinct storage mechanisms of soil organic carbon in coniferous forest and evergreen broadleaf forest in tropical China[J]. Journal of Environmental Management, 295: 113142. doi: 10.1016/j.jenvman.2021.113142
Sun T, Wang Y, Hui D, et al. 2020. Soil properties rather than climate and ecosystem type control the vertical variations of soil organic carbon, microbial carbon, and microbial quotient[J]. Soil Biology and Biochemistry, 32: 106147.
Taylor L A, Arthur M A, Yanai R D. 1999. Forest floor microbial biomass across a northern hardwood successional sequence[J]. Soil Biology and Biochemistry, 31(3): 431−439. doi: 10.1016/S0038-0717(98)00148-5
Vance E D, Brookes P C, Jenkinson D S. 1987. An extraction method for measuring soil microbial biomass C[J]. Soil Biology and Biochemistry, 19(6): 703−707.
Wang Q C, Zhen Y, Song G, et al. 2021. Impacts of simulated nitrogen and phosphorus depositions on soil microbial biomass and soil nutrients along two secondary succession stages in a subtropical forest[J]. Acta Ecologica Sinica, 41(15): 6245−6256 (in Chinese with English abstract).
Wang S Q, Yu G R. 2008. Ecological stoichiometry characteristics of ecosystem carbon, nitrogen and phosphorus elements[J]. Acta Ecologica Sinica, 28(8): 3938−3945 (in Chinese with English abstract).
Wuhan Municipal Landscape Gardens and Forestry Bureau, Annual Report on Greening of Wuhan City in 2021[EB/OL]. (2022−03−17). [2022−12−07]. http://ylj.wuhan.gov.cn/zwgk/zwxxgkzl_12298/tjxx/lhgb_12361/202203/t20220317_1941383.shtml (in Chinese).
Xu X, Han L, Wang Y, et al. 2007. Influence of vegetation types and soil properties on microbial biomass carbon and metabolic quotients in temperate volcanic and tropical forest soils[J]. Soil Science and Plant Nutrition, 253: 430−440.
Yang J H, Wang C L, Dai H L. 2008. Soil agrochemical analysis and environmental monitoring[M]. Beijing: China Land Publishing House (in Chinese).
You Y M, Wang J, Huang X M, et al. 2016. Responses of soil microbial biomass, community structure and soil enzyme to below−ground carbon changein the warm−temperate forest ecosystem[J]. Guihaia, 36(7): 837−847 (in Chinese with English abstract).
Zhang H Y. 2005. A study on the relationship between black soil microbiomass and soil fertility[D]. Master Thesis of Shenyang Agricultural University (in Chinese with English abstract).
Zhang Y Q, Fang X, Xian Y N, et al. 2019. Characteristics of soil microbial biomass carbon, nitrogen, phosphorus and enzyme activity in four subtropical forests, China[J]. Acta Ecologica Sinica, 39(14): 5326−5338 (in Chinese with English abstract).
陈闽昆, 王邵军, 陈武强, 等. 2019. 蚂蚁筑巢对西双版纳热带森林土壤微生物生物量碳及熵的影响[J]. 应用生态学报, 30: 2973−2982. 陈小花, 陈宗铸, 雷金睿, 等. 2021. 东寨港不同植物群落土壤微生物量碳氮及养分特征[J]. 林业资源管理, (6): 97−104. 程谊, 张金波, 蔡祖聪. 2012. 土壤中无机氮的微生物同化和非生物固定作用研究进展[J]. 土壤学报, 49(5): 1030−1036. 丁令智, 满秀玲, 肖瑞晗, 等. 2019. 寒温带森林根际土壤微生物量碳氮含量生长季内动态变化[J]. 林业科学, 55(7): 178−186. 高珊, 伊航, 傅民杰, 等. 2018. 冻融循环对温带3种林型下土壤微生物量碳、氮和氮矿化的影响[J]. 生态学报, 38(21): 7859−7869. 管海英, 王权, 赵鑫, 等. 2015. 两种典型荒漠植被区土壤微生物量碳的季节变化及影响因素分析[J]. 干旱区地理, 38(1): 67−74. 胡宗达, 刘世荣, 史作民, 等. 2012. 川滇高山栎林土壤氮素和微生物量碳氮随海拔变化的特征[J]. 林业科学研究, 25(3): 261−268. 黎荣彬. 2008. 土壤微生物生物量碳研究进展[J]. 林业与环境科学, 24: 65−68. doi: 10.3969/j.issn.1006-4427.2008.02.015 李雅, 卢杰. 2023. 土壤碳的影响因素及凋落物对土壤碳储量的影响[J]. 农业与技术, 43(4): 52−55. 刘爽. 2010. 五种温带森林土壤微生物生物量碳和氮的时空变化[D]. 东北林业大学硕士学位论文. 马彩霞, 李洪杰, 郑海峰, 等. 2019. 川西高山林线土壤活性碳 氮对短期增温的响应[J]. 应用生态学报, 30(3): 718−726. 青烨, 孙飞达, 李勇, 等. 2015. 若尔盖高寒退化湿地土壤碳氮磷比及相关性分析[J]. 草业学报, 24(3): 38−47. doi: 10.11686/cyxb20150304 全国土壤普查办公室. 1979. 全国第二次土壤普查暂行技术规程[M]. 农业出版社出版: 1−79. 施福军, 黄则月, 李婷, 等. 2018. 望天树天然林土壤微生物生物量碳氮垂直分布及相关性分析[J]. 林业与环境科学, 34: 72−76. doi: 10.3969/j.issn.1006-4427.2018.06.012 王全成, 郑勇, 宋鸽, 等. 2021. 亚热带次级森林演替过程中模拟氮磷沉降对土壤微生物生物量及土壤养分的影响[J]. 生态学报, 41(15): 6245−6256. 王绍强, 于贵瑞. 2008. 生态系统碳氮磷元素的生态化学计量学特征[J]. 生态学报, 28(8): 3938−3945. doi: 10.3321/j.issn:1000-0933.2008.08.054 武汉市园林和林业局. 2021年武汉市绿化状况公报[EB/OL]. (2022−03−17). [2022−12−07]. http://ylj.wuhan.gov.cn/zwgk/zwxxgkzl_12298/tjxx/lhgb_12361/202203/t20220317_1941383.shtml. 杨剑虹, 王成林, 代亨林. 2008. 土壤农化分析与环境监测[M]. 北京: 中国大地出版社. 尤业明, 王娟, 黄雪蔓, 等. 2016. 暖温带森林土壤微生物量、群落结构和活性对植物地下碳源的响应[J]. 广西植物, 36(7): 837−847. doi: 10.11931/guihaia.gxzw201409049 张海燕. 2005. 有关黑土微生物量与土壤肥力关系的研究[D]. 沈阳农业大学硕士学位论文. 张雅茜, 方晞, 冼应男, 等. 2019. 亚热带区4种林地土壤微生物生物量碳氮磷及酶活性特征[J]. 生态学报, 39(14): 5326−5338.






计量
- 文章访问数: 24687
- HTML全文浏览量: 265
- PDF下载量: 317
